Fatty acids and astaxanthin composition of two edible native Mexican crayfish Cambarellus (C.) montezumae and Procambarus (M.) bouvieri
G. Coral-Hinostrozaa,*, M. Díaz-Martíneza, A. Hubermanb and J.L. Silencio-Barritac
aDepartment of Animal Nutrition, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Vasco de Quiroga 15, 14080 D.F. Mexico
bDepartment of Biochemistry, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Vasco de Quiroga 15, 14080 D.F. Mexico
cDepartment of Science and Food Technology, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Vasco de Quiroga 15, 14080 D.F. Mexico
*Corresponding author: gldscoral9@gmail.com
| |
SUMMARY
The content and composition of the fatty acids (FAs) and astaxanthin (AST) in the edible forms of crayfish: the whole animal
of Cambarellus (C.) montezumae, and the tail meat (TM) of Procambarus (M.) bouvieri were determined by GC and HPLC. The exoskeleton (EXK) of P. (M.) bouvieri was also studied. Unsaturated FAs, and mostly oleic acid (C18:1 n-9), were predominant in both edible forms. The contents
of the polyunsaturated eicosapentaenoic (C20:5 n-3, EPA), arachidonic (C20:4 n-6, ARA) and docosahexaenoic acid (C22:6 n-3,
DHA), were higher in the TM of P. (M.) bouvieri than in the complete C. (C.) montezumae (p<0.05). Total carotenoids ranged between 2.31 ± 0.33 μg·g−1 and 66.3 ± 3.91 μg·g−1, and were composed mainly of AST (>79.50%). AST esters were enriched with saturated FAs in C. (C.) montezumae and with PUFAs in EXK of P. (M.) bouvieri. We conclude that both C. (C.) montezumae and the TM of P. (M.) bouvieri are traditional foods rich in n-3 PUFAs and C. (C.) montezumae in AST. The EXK of P. (M.) bouvieri is a rich potential source of AST, n-3 PUFAs, and the combination AST-DHA.
|
| |
RESUMEN
Composición de ácidos grasos y astaxantina de dos especies comestibles de acociles nativos de México, Cambarellus (C.) montezumae y Procambarus (M.) bouvieri. Se determinó por GC y HPLC el contenido y composición de ácidos grasos (AGs) y astaxantina (AST), en dos formas comestibles
de acocil: el animal completo de Cambarellus (C.) montezumae, y el músculo de la cola (MC) de Procambarus (M.) bouvieri. Adicionalmente, se estudió el exosqueleto (EXK) de P. (M.) bouvieri. En ambas formas comestibles predominaron los AGs insaturados. Los contenidos de ácido eicosapentaenoico (C20:5 n-3, EPA),
araquidónico (C20:4 n-6, ARA) y docosahexaenoico (C22: 6 n-3, DHA), fueron mayores en el MC que en C. (C) montezumae (p <0,05). Los carotenoides totales oscilaron de 2.3 ± 0.3 μg·g−1 a 66.3 ± 3.9 μg·g−1, con predominancia de AST (>79.50%). Los ésteres de AST en C. (C.) montezumae fueron enriquecidos con AGs saturados mientras que los del EXK de P. (M.) bouvieri con AGs poliinsaturados. Se concluyó que tanto C. (C.) montezumae como el MC de P. (M.) bouvieri, son alimentos tradicionales ricos en PUFAs n-3, y C. (C.) montezumae en AST. El EXK de P. (M.) bouvieri abunda en AST, PUFAs n-3, y en la combinación AST-DHA.
|
1. INTRODUCTIONTOP
Wild crayfish have been caught and consumed by native Mexicans since pre-Hispanic times. Cambarellus (Cambarellus) montezumae (de Saussure, 1857), (C. (C.) montezumae), and Procambarus (Mexicambarus) bouvieri (Ortman, 1909), (P. (M.) bouvieri), are two autochthonous species among the 56 crayfish of the Cambaridae family that inhabit the fresh waters of Mexico (Alvarez et al., 2012). Commonly known as “acocil”, derived from the Nahuatl language “acotzilli”, they are ingredients of traditional food. C. (C.) montezumae, is the most widely distributed species along the neo-volcanic axis of Mexico (Alvarez et al., 2012). This organism has a small average adult size (32±4.3 mm total length), with a soft and thin EXK so that the consumption
of the entire animal is common, either boiled, fried, or roasted. The P. (M.) bouvieri is found in Uruapan, Michoacan, Mexico. It is larger than C. (C.) montezumae (with an average adult size of 70.5±8.1 mm total length). As in other edible crayfish only the TM is consumed. When whole
crayfish are boiled in a soup, the TM is eaten and the EXK is discarded. Both species are caught by the natives and are not
marketed. At present, there is no crayfish aquaculture in Mexico, although some prospective studies consider its feasibility
(Cerón-Ortiz et al., 2015).
Crayfish lipids, as in other crustaceans, are important components characterized mainly by their high phospholipid (PL) content
and essential fatty acids (EFA), the polyunsaturated (PUFAs) linoleic acid (C18:2n-6, LA), α-linolenic acid (C18:3n-3, ALA),
and chiefly the readily available essential eicosapentaenoic (C20:5n-3, EPA), and docosahexaenoic acid (C22:6n-3, DHA) (Stanek
et al., 2011; Harlioğlu et al., 2012; Harlioğlu et al., 2015). There is a current criterion about the beneficial effect of the intake of n-3 PUFAs to prevent many degenerative diseases.
To this end, scientific international committee have recommended a dietary intake of 250 mg per day of EPA+DHA (FAO, 2010). Along with FAs, the carotenoid astaxanthin (AST) (3,3′-dihydroxy-β,β’-carotene-4,4′-dione), is found in different crustacean
tissues in free and esterified forms (Sachindra et al., 2005). The physiological functions of this fat-soluble antioxidant have been widely documented in vitro, in vivo, and in pre-clinical studies and covered extensively for its health benefits in a recent review (Dose et al., 2016). AST associated to EPA and DHA has been demonstrated to have a great effectiveness for improving human brain function (Konagai
et al., 2013), and neuro-protection (Barros et al., 2014). In this respect, Saw et al. (2013), confirmed that low doses of AST combined with DHA or EPA have a synergistic antioxidant action mediated by the Nrf2 signaling
pathway.
The fatty acids and carotenoids in crayfish have been sparsely investigated compared to marine crustaceans. The North American
species Astacus leptodactylus (Harlioğlu et al., 2012; Harlioğlu et al., 2015), and Procambarus clarkii (Meyers and Bligh, 1981); the European Astacus astacus (Ackefors et al., 1997), and the Australian Cherax quadricarinatus (Sagi et al., 1995; Harlioğlu et al., 2012), have been the most investigated in the world so far. Particularly, C. (C.) montezumae post-larvae has recently been studied regarding the FA composition (Cerón-Ortiz et al., 2015), but not about carotenoid content while P. (M.) bouvieri has not yet been studied with regards to these topics. Thus, the aim of the present study was to determine the content and
composition of FAs and AST in the two edible forms of these crayfish species: the whole animal of C. (C.) montezumae, and the TM of P. (M.) bouvieri. In addition, the study was extended to the EXK of P. (M.) bouvieri as a potential source of these important molecules.
2. MATERIAL AND METHODSTOP
2.1. Reagents and materialsTOP
Organic solvents were of analytical grade and, when necessary, of HPLC grade, obtained from the Sigma-Aldrich Corporation
(St Louis, MO, USA). Synthetic AST was obtained from Hoffmann-La Roche (Basel, Switzerland). β-carotene (purity: 97%), and
fatty acid methyl esters (FAME) standards were purchased from Sigma Chemical Co. (St. Louis, MO, USA). All other reagents
used were of analytical grade.
2.2. EquipmentTOP
Capillary gas chromatography (GC) was carried out in a CP-3380 gas chromatograph equipped with a flame ionization detector
and a CP-8400 autosampler (Varian Inc., Palo Alto, CA, USA). HPLC analysis was performed on a HPLC Waters system with a 717
autosampler, 1525 binary pump, and a dual 2487 UV/VIS absorbance detector (Milford, MA, USA). Sample absorbances were measured
in a DU-60 Beckman spectrophotometer (Beckman, Fullerton, CA, USA). Vacuum evaporation was carried out on a rotary evaporator
(Rotavapor R-134, Büchi, Brinkmann Instruments, Inc., N. Y., USA).
2.3. Animals and sample preparationTOP
Adult C. (C.) montezumae of both sexes (with a total length size >30 mm), were caught in June in the nearby canals of Xochimilco, Mexico City. They
were transported to the laboratory alive in three polyethylene bags with water, and placed over crushed ice (4 °C). Then,
the bags were drained, wrapped with Aluminium foil and frozen at –80 °C. As these animals are consumed entire without sex
selection, the experimental sample consisted of a mixed capture. After defrosting, three kilograms of dead animals were washed
with tap water and left to dry in a pilot plant cabinet dryer (Apex, model SSE 17M) at 50 ± 2 °C for 24 h, with natural air
circulation. The dried sample was ground in a Wiley model 3379-K41 knife mill (Thomas Scientific, Philadelphia, PA, USA) using
a 2 mm mesh. The meal was kept in a polyethylene bag at –20 °C until analysis. Adult P. (M.) bouvieri (total length size >70 mm), were caught in June in an Uruapan stream, Michoacan, transported to the laboratory alive in a
cooler with crushed ice (4°C) and stored in plastic containers at –80 °C until analysis. After thawing, eighty dead animals
with a mean individual body weight of 17.4 ± 0.6 g, were separated into the TM portion (14.7±2.5%, mean yield of the body
weight), and EXK. The TM was analyzed fresh. EXK, composed of cephalothorax and abdomen shell, was manually and carefully
freed from soft tissue, washed under running water, dried and processed to a meal as described above. The final moisture content
of C. (C.) montezumae meal was 18.5 ± 1.3% and that of EXK was 12.2 ± 0.9%. The resulting meals were split into three parts and each analyzed in
triplicate. In both species the animals were caught and processed in bulk, without sex separation.
2.4. Preparation of oil extractsTOP
Meal samples of C. (C.) montezumae, the TM and the EXK of P. (M.) bouvieri, were subjected to analytical procedures under reduced white light. A 20 g meal sample from each species, or 50 g TM (wet
weight 75.4 ± 3.5 g·100 g−1) were ground in a mortar and extracted repeatedly with cold acetone (4 °C) until no further pigment was extractable. The
colored acetone extracts were pooled and filtered through a Whatman No.1 filter paper under vacuum, phase-separated in 50
mL of petroleum ether (PE) in a separating funnel of 250 mL and washing repeatedly with water to remove traces of acetone.
The organic phase was then filtered through a coarse-sintered glass funnel containing anhydrous sodium sulfate, rinsing the
pigments with additional PE. Finally, the solvent was evaporated to dryness in a rotary evaporator at 45 °C under vacuum to
obtain the oil extract which was stored in the dark under nitrogen at –20 °C until analysis.
2.5. Determination of fatty acidsTOP
Fatty acids in the oil extracts of all the samples were converted into methyl esters with 14% methanolic boron trifluoride,
according to the AOAC method 969.33 (AOAC, 2000). Chromatographic separation was performed with a DB-23 capillary column (length 30 m, i. d. 0.25 mm, film thickness 25 μm)
(J and W Scientific, Folsom, CA, USA). Nitrogen was used as carrier gas at a flow rate of 2 mL·min−1. Oven temperature was programmed for a linear increase of 10 °C·min−1 from 120 °C to 200 °C and 5 °C·min−1 from 200 °C to 230 °C. Myristoleic acid (C14:1) (Sigma-Aldrich), was used as internal standard. The chromatographic peaks
were identified by comparison of their retention times with those of standards and quantified with the Star Workstation program
(Varian Inc.), using a mixture of pure standards to detect retention times. Each identified FA was expressed as a percentage
of the total FAs recorded in the sample. Additionally, the quality of the oil extracts was calculated by the formulae of the
atherogenicity index (AI) and thrombogenicity index (TI), proposed by Ulbricht and Southgate, (1991):
AI = [ (C12:0 + (4 x C14:0) + C16:0) ] / [ MUFA + (PUFA n-6 + n-3) ] (Eq. 1)
TI = [C14:0+C16:0 + C18:0] / [(0.5 x MUFA) + (0.5 x PUFA n-6) + (3 x PUFAn-3) + PUFA n-3/PUFAn-6] (Eq. 2)
2.6. Total carotenoid quantificationTOP
Oil extracts from each replicated extraction of samples were taken in a known volume of n-hexane and the carotenoids were quantified by determining the optical density at 470 nm (absorption maximum of AST in n-hexane). Since AST is the main carotenoid in crustaceans, the total concentration of carotenoids was referred to as this
carotenoid in the oil extracts, according to Sachindra et al. (2005), by means of the formula:
Carotenoid content (μg AST · g−1 sample) = A × V × D/0.2 × W (Eq. 3)
where, A = absorbance at 470 nm; V = volume of the extract in n-hexane; D = dilution factor; 0.2 is the absorbance value (λ = 470 nm) of 1 μg·mL−1 AST standard in n-hexane, and W is the weight of meal or TM sample (g).
The absorption maximum and chromatographic features of the AST standard were obtained. To this end, 1.5 mg of synthetic compound
were dissolved in 5 mL chloroform and brought up to 50 mL with n-hexane (stock solution). A standard solution of AST was made from the stock solution in n-hexane:chloroform (95.5:4.5, v/v), and absorption spectra were determined between 360 and 600 nm.
2.7. Chromatographic separation of carotenoidsTOP
Individual carotenoids in total oil extracts were separated, identified and their relative composition was established by
chromatography, except those of TM, whose separation was not possible due to insufficient quantity.
2.7.1. Open column chromatography (OCC)TOP
Two g of oil extract diluted in 25 mL n-hexane were eluted from a silica gel column 32 × 420 mm (Si 60, particle size 0.40–0.63 mm), with stepwise 5% decreasing
proportions of n-hexane in acetone, following the procedure described by Coral-Hinostroza and Bjerkeng (2002). The yellow non-polar lipids (fraction F1), were first eluted with pure n-hexane; the next two orange-red colored F2 and F3 were separated with a mixture of n-hexane:acetone elution solvent in a proportion of 95:5 (v/v), and 90:10 (v/v), respectively; and the last red fraction (F4)
with 85:15 (v/v). To increase the purity of the F2, F3, and F4 fractions, they were re-chromatographed with the same mixture
of solvents starting with pure n-hexane. The obtained fractions were evaporated to dryness, re-dissolved in HPLC grade n-hexane for TLC and HPLC tests, and subjected to spectroscopy for obtaining their absorption spectra (360 to 600 nm) and maximum
wavelengths (λmax).
2.7.2. TLC ChromatographyTOP
Fractions F2, F3, and F4 were applied individually to Aluminium backed plates pre-coated with silica (0.2 mm, Merck DC-alufolien,
Kieselgel 60 F254, Darmstadt, Germany), and their retention factor (Rf) was calculated in parallel with the synthetic AST, AST-monoester and AST-diester standards. The last two were prepared by
separation and purification from the red crab Pleuroncodes planipes (Coral-Hinostroza and Bjerkeng, 2002). Spots were eluted in a mobile phase of n-hexane:acetone (70:30, v/v).
2.7.3. HPLC chromatographyTOP
Carotenoids were analyzed by HPLC on a H3PO4-modified Spherisorb SW column (250 mm × 4.6 mm i.d., 5 μm, Waters) at a controlled temperature (30 °C) with a mobile phase
of n-hexane: acetone (86:14, v/v). The flow rate and injection volume were 1.0 mL min−1 and 20 μL, respectively, and the detection wavelength was set at 470 nm. Fractions F2, F3, and F4, were filtered with a pore
size 0.45 μm PVLP membrane Durapore filter (Millipore, Bedford, MA, USA), and injected. The HPLC spectra of the synthetic AST
and the prepared AST-ester standards were obtained under the same chromatographic conditions as the samples. Carotenoid separation
of the complete oil extracts was also obtained by dissolving them (1:100, v/v) in HPLC grade n-hexane, through a 0.45 μm pore size PVLP Durapore membrane (Millipore, Bedford, MA, USA), and eluting for 15 min under the
same conditions as the standards. The identification of carotenoids was based on the comparison of the retention times of
sample peaks with those of standards. The relative content of total AST was obtained by summing up the areas under the curves
of monoester, diester, and free AST, considering the total carotenoid content as 100%. The content of free AST was calculated
by summing up the areas under the curves of its geometrical isomers.
2.8. Qualitative analysis of fatty acid composition of the AST estersTOP
Qualitative analysis of FA composition in AST-monoester, and AST-diester fractions obtained from the OCC separation were undertaken
using gas chromatography after the methylation step following the procedure explained above.
2.9. Statistical analysesTOP
Results are expressed as mean values of three determinations ± standard deviation (SD). An analysis of variance (ANOVA) was
carried out to determine the difference among the examined group of samples. A comparison of mean values was carried out by
Duncan's multiple range test and statistically significant differences were reported at p < 0.05. Data analyses were performed using the Statistical Package for Social Science (SPSS 11.0 for Windows; SPSS, Chicago,
IL, USA).
3. RESULTS AND DISCUSSIONTOP
3.1. Fatty acids of oil extractsTOP
The FA profiles of the oil extracts from C. (C.) montezumae, and the TM of P. (M.) bouvieri are summarized in Table 1. Palmitic acid (C16:0) and oleic acid (C18:1n-9) were the prevailing saturated (SFA) and monounsaturated fatty acids (MUFA),
in both analyzed samples, whereas EPA and LA were the dominant PUFAs. SFA pentadecanoic (C15:0) was found only in the TM of
P. (M.) bouvieri. The general composition pattern of FAs in both edible forms are similar to those found in complete C. (C.) montezumae (Cerón et al., 2015), and to other crayfish species (Stanek et al., 2011; Harlioğlu et al., 2012; Harlioğlu et al., 2015). Mean percentage of SFAs (26.52 ± 0.41%) and MUFAs (42.80 ± 0.57%), were significantly higher in C. (C.) montezumae than in the TM of P. (M.) bouvieri (SFAs, 24.04 ± 0.15% and MUFAs, 34.22 ± 0.82%) (p<0.05). The high levels of SFAs and MUFAs in the entire animals of C. (C.) montezumae were probably derived from organs, such as the hepatopancreas and gonads of possible female animals in the sample, of high
lipid content, mainly C18:1n-9, which is stored as an energy source (Gonzalez-Baro and Pollero, 1988).
Table 1. Fatty acid profile and lipid quality indices of the oil extracts of the two crayfish species, Cambarellus (C.) montezumae and Procambarus (M.) bouvieri, expressed as percentage (%) of the total fatty acids
| Fatty acid |
Edible forms |
P. (M.) bouvieri EXK
|
| C. (C.) montezumae whole animal
|
P. (M.) bouvieri TM
|
| SFA |
|
|
|
| C14:0 |
|
|
1.37±0.31 |
| C15:0 |
|
0.35±0.05a |
0.93±0.15b |
| C16:0 |
20.46±1.02c |
17.20±0.60a |
18.31±0.25b |
| C18:0 |
5.01±0.35a |
5.78±0.64a |
5.52±0.33a |
| C20:0 |
1.05±0.28b |
0.70±0.06a |
0.82±0.07a |
| MUFA |
|
|
|
| C14:1n-7 |
10.45±1.69c |
5.27±0.50a |
7.67±0.40b |
| C16:1n-7 |
10.60±0.43b |
8.36±0.23a |
10.08±0.43b |
| C17:1n-8 |
|
|
1.61±0.07 |
| C18:1n-9 |
21.74±1.11b |
20.60±0.12b |
18.38±0.23a |
| PUFA |
|
|
|
| C18:2n-6 |
9.31±0.16a |
9.10±0.40a |
10.25±0.11b |
| C18:3n-3 |
4.15±0.88b |
3.56±0.08a |
5.19±0.17b |
| C20:4n-6 |
4.14±0.45a |
9.83±0.45c |
5.31±0.06b |
| C20:5n-3 |
10.35±0.59a |
13.39±0.15c |
11.51±0.26b |
| C22:6n-3 |
2.19±0.30a |
5.32±0.29c |
3.02±0.16b |
| ΣSFA |
26.52±0.41b |
24.04±0.15a |
26.95±0.35b |
| ΣMUFA |
42.80±0.57c |
34.22±0.82a |
37.74±0.30b |
| ΣPUFA |
30.14±1.30a |
41.19±0.88c |
35.28±0.18b |
| Total n-3 |
16.99±1.09a |
22.26±0.11c |
19.72±0.19b |
| Total n-6 |
13.45±0.38a |
18.93±0.37c |
15.55±0.11b |
| n-3/n-6 |
1.26±0.03b |
1.18±0.04a |
1.27±0.01b |
| AI |
0.28±0.01b |
0.23±0.01a |
0.33±0.01c |
| TI |
0.32±0.01c |
0.24±0.00a |
0.29±0.01b |
| TM, tail meat; EXK, exoskeleton; AI, atherogenicity index; TI, thrombogenicity index. |
| Means with different superscripts in the same row are significantly different (p < 0.05)
|
Total PUFAs, were significantly greater in TM (41.19 ± 0.88%) than in C. (C.) montezumae (30.14 ± 1.30%) (p<0.05), with statistically significant predominance of EPA, arachidonic acid (C20:4n-6, ARA), and DHA in the former than in
the latter. The discrepancy in the FA content and composition between our samples might be attributed to:1) different animal
components in the analyzed sample, and 2) to different habitat conditions of each species, in particular diet composition,
in line with Cerón-Ortiz et al. (2015). The contents of LA in our studied crayfish were high, contrasting with those of seafood that are characterized by low levels
of n-6 and high levels of n-3 (Valfré et al., 2003). There is evidence that EPA and DHA in the PL form ingested directly possess high bioavailability in humans (Konagai et al., 2013; Cook et al., 2016), and that consumption of krill oil increases these long chain EFA concentrations in plasma and blood cells (Ramprasath et al., 2013). The n-3/n-6 ratio values for C. (C.) montezumae (1.26 ± 0.03) was of higher magnitude than that of the TM (1.18±0.04) (p<0.05). Lower ratios were found in TM of wild crayfish Orconectes limosus, aged 4+ (Stanek et al., 2011), while higher values were determined in the TM of wild A. leptodactylus (Harlioğlu et al., 2012). The ratio value for C. (C.) montezumae was comparable to those observed in the TM of A. leptodactylus (both sexes), after a diet supplementation with 1% n-3 PUFAs (Harlioğlu et al., 2015). Our obtained values were in the range of 1 to 4 as observed for fish meat (Valfré et al., 2003), suggesting the good quality of these crayfish species as a food.
Ulbricht and Southgate (1991) have indicated that fats with high atherogenicity (AI), and thrombogenicity (TI) indices are assumed to be detrimental to
human health. As shown in Table 1, the AI of whole C. (C.) montezumae oil extracts were 0.28 ± 0.01 and in the TM of P. (M.) bouvieri 0.23 ± 0.01. These values are lower compared to those reported for rainbow trout meat (0.57) (Valfré et al., 2003), sardine (0.60), and mackerel (0.48) (Fernandes et al., 2014), and much lower than that of terrestrial animal meat such as lamb (1.00), beef (0.72), pork (0.69), chicken (0.50), and
rabbit (0.82) (Bosco et al., 2001). Mean values of TI were 0.24 ± 0.00 and 0.32 ± 0.01 in the TM of P. (M.) bouvieri and C. (C.) montezumae, respectively, which are in the range of 0.20 to 0.40 just as those found for marine fish meat (Fernandes et al., 2014). From our results, we can suggest that the utilization of both these edible forms of crayfish species for human nutrition
offer advantages because they have a good n-3/n-6 balance and lipids of good quality, in the context of the low intake of
n-3 PUFAs in the Mexican population as claimed by Ramirez-Silva et al. (2011), derived probably from the patterns of food choice and unavailability of fish stocks to supply fish meat.
As shown in Table 1, the two most important FAs by order of magnitude in the EXK of P. (M.) bouvieri were C18:1n-9 and C16:0 as was observed in the edible samples. In addition, three different FAs were identified, myristic
(C14:0), pentadecanoic (C15:0) and margaroleic (C17:1n-8), although in minimal amounts (<1.6% each). Among the PUFAs, high
amounts of EPA and LA where determined in this crayfish component. The n-3/n-6 ratio in it was 1.27 ± 0.01, whereas values
of TI (0.29) and AI (0.33) were in the range of those reported for some marine fish (Fernandes et al., 2014). This pattern, associated with a higher n-3 content in the oil extract of EXK of P. (M.) bouvieri contributes to a desirable nutritional quality with high potential as a valuable ingredient in the formulation of animal
feed.
3.2. Carotenoid content and composition in the oil extractsTOP
The total carotenoid concentration in the whole C. (C.) montezumae was 66.30 ± 3.95 (μg AST·g−1), significantly higher than that in the TM of P. (M.) bouvieri (2.30 ± 0.29 μg AST·g−1) (p < 0.05) (Table 2). The carotenoid amount in the former, measured as AST, is notably higher than that determined in a meal of whole Procambarus clarkii (15.74 μg·g−1) (García-Romero et al., 2014). The high carotenoid level in a whole crayfish can be anticipated as it includes organs rich in AST such as the gonads of
ovigerous females, eye stalks, antenna, and components such as the EXK where high levels of this pigment compound is deposited
as demonstrated in A. leptodactylus (Czeczuga, 1971). On the contrary, the carotenoid content in the muscle tissue of these animals is low as affirmed by Czeczuga (1971), in A. leptodactylus (1.3 μg g−1) and in Cherax quadricarinatus (0.04 to 0.62 μg·g−1) (Sagi et al., 1995). Varying AST concentrations in tissue may be attributed to the available dietary carotenoids of crayfish in their particular
environments or to the metabolic needs of particular carotenoid in the organ. The present study has shown the high pigment
importance of a C. (C.) montezumae meal, with concentrations as much as 2.6 times greater than that reported for the meal of sun-dried shrimp, Litopenaeus vannamei (25.5 μg·g−1) (Hernandez et al., 2014), another edible form of a complete crustacean consumed in Mexico.
Table 2. Total carotenoid content in the two edible forms of crayfish Cambarellus (C.) montezumae and Procambarus (M.) bouvieri; and in the exoskeleton of Procambarus (M.) bouvieri
| Total carotenoid content |
Edible forms |
P. (M.) bouvieri EXK
|
| C. (C.) montezumae whole animal
|
P. (M.? bouvieri TM
|
| μg AST·g−1 |
66.30 ± 3.95b |
2.30 ± 0.29a |
424.40 ± 9.02c |
| TM, tail meat; EXK, exoskeleton. Data expressed as mean ± standard deviation (SD). Different superscripts in the row are significantly
different (p < 0.05).
|
The EXK of P. (M.) bouvieri showed a high content of carotenoids (424.40 ± 9.02 μgAST·g−1) with respect to the edible forms of this study. This content was lower than the one found in the purified carotenoprotein-1
fraction of Procambarus clarkii (610 μg·g−1) (Cremades et al., 2003). As noted by Zagalsky et al. (1990), AST is incorporated as the prosthetic group into proteins of crustacean shells in which it is mostly deposited and are
responsible for coloration. It could then be expected that animals with intense colors provide significant levels of this
carotenoid as is the case of P. (M.) bouvieri which when alive shows an external dark orange-brown color.
The estimation of the carotenoid composition by OCC resulted in four fractions. F1, β-carotene; F2, diester of AST; F3, monoesters
of AST; and F4, free AST with their chromatographic features presented in Table 3. The HPLC profile of the complete oil extracts (Figure 1) of C. (C.) montezumae (Chromatogram A) and EXK (Chromatogram B), showed that AST (peak 3) and its esters (peaks 1, 2) were the major carotenoid
pigments in both samples (Table 4). In addition to AST, lesser amounts of β-carotene and other not identified compounds were distinguished, more notably in
C. (C.) montezumae than in EXK, which may correspond to the influence of carotenoids from organs such as the hepatopancreas, whose principal
carotenoid is β-carotene (Sagi et al., 1995). The ocurrence of AST as the main carotenoid in species of the current study was congruent with those found in other crayfish
species such as Procambarus clarki and Astacus astacus, as mentioned above.
|
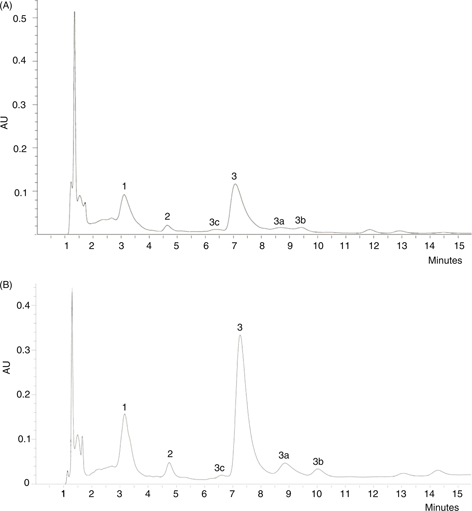 Figure 1. HPLC comparison of astaxanthin (AST) composition in the whole crayfish Cambarellus (C.) montezumae (A), and Procambarus (M.) bouvieri exosqueleton (B). Peaks were identified by retention time, co-chromatography with synthetic AST and AST-esters. Peak 1: AST-diester;
peak 2: AST-monoester; peak 3: trans all-E AST showing its geometrical isomers: 3a, 9 cis; 3b, 13 cis; 3c, probably Di-Z. Figure 1. HPLC comparison of astaxanthin (AST) composition in the whole crayfish Cambarellus (C.) montezumae (A), and Procambarus (M.) bouvieri exosqueleton (B). Peaks were identified by retention time, co-chromatography with synthetic AST and AST-esters. Peak 1: AST-diester;
peak 2: AST-monoester; peak 3: trans all-E AST showing its geometrical isomers: 3a, 9 cis; 3b, 13 cis; 3c, probably Di-Z.
|
|
Table 3. Chromatographic and UV/VIS features of forms of astaxanthin in the oil extract samples of Cambarellus (C.) montezumae and Procambarus (M.) bouvieri crayfish
| Fraction |
OCC elution mixture (n-hexane:acetone, (v/v)
|
UV-VIS λ max (n-hexane) (nm)
|
TLC (Rf) solvent system (n-hexane:acetone) (70:30, v/v)
|
| F1, β-carotene |
100 |
449 |
0.98 ± 0.02 |
| F2, di-AST |
95:5 |
468 |
0.94 ± 0.01 |
| F3, mono-AST |
90:10 |
469 |
0.70 ± 0.02 |
| F4, free AST |
85:15 |
470 |
0.33 ± 0.01 |
| F2, diester of AST; F3, monoester of AST; OCC, open column chromatography; Rf, retention factor value; UV/VIS, maximum absorption
in n-hexane.
|
Table 4. Relative composition of major carotenoids in the whole Cambarellus (C.) montezumae and in the exoskeleton of Procambarus (M.) bouvieri by HPLC
| Peak |
Carotenoids |
C. (C.) montezumae whole animal
|
P. (M.) bouvieri EXK
|
|
Total AST |
79.50±1.02a |
93.15±5.71b |
|
β-carotene |
15.48±2.60b |
4.85±5.44a |
|
NI |
5.02±3.15b |
2.00±0.27a |
|
|
|
|
|
AST fractions* |
|
|
| 1 |
di-AST |
54.39±1.02b |
35.07±3.00a |
| 2 |
Mono-AST |
3.73±0.15a |
3.81 ±1.56a |
|
Free AST |
41.88±1.17a |
61.13±1.44b |
|
|
|
|
|
AST isomers** |
|
|
| 3c |
Di-Z |
1.94±0.16a |
1.45±0.35a |
| 3 |
all-trans |
81.64±0.85a |
82.71± 0.42a |
| 3a |
9-cis |
8.61± 0.64b |
6.02± 0.82a |
| 3b |
13-cis |
7.93± 0.44a |
9.81± 0.17b |
| EXK, exoskeleton; Peaks 1, 2, 3c, 3, 3a, and 3b, correspond to Figure 1.
|
| (*) Percentages calculated with respect to total AST. |
| (**) Percentage of isomers calculated with respect to free AST. NI: not identified carotenoids. Data were expressed as mean
± standard deviation (SD). Different superscripts in the same row are significantly different (p < 0.05).
|
The relative AST content in the EXK of P. (M.) bouvieri was 93.15 ± 5.71%, significantly higher than that of C. (C.) montezumae (79.50 ± 1.02%) (p < 0.05) (Table 4). Both samples were distinguished by their high content of free AST (61.13 ± 1.44% and 41.88 ± 1.17%, respectively), although
the content in EXK surpassed 50% with respect to the esterified forms. A high content of free AST was observed earlier in
the waste products of Procambarus clarki (40.3% free and 49.4% esterified) (Meyers and Blight, 1981), and in the EXK of the same species where free AST was the main form (Inoue et al., 1988). It seems that in Procambarus crayfish species the prevailing form of this carotenoid is free and might be attributable to a particular biochemical characteristic.
Taking into account the carotenoid concentration of the whole C. (C.) montezumae, and its relative AST content it is worth considering the consumption of this food as a good way to enrich the diet with
this bioactive molecule, although its bioavailability in these edible forms remains to be investigated. The geometric isomer
composition of free AST in all cases showed a higher proportion of trans all-E isomer (around 80%), and minor amounts of 9cis and 13cis configurations (around 15%), which corresponds to the normal composition of this carotenoid in nature. Di-Z configuration could not be confirmed.
3.3. Fatty acid profile of astaxanthin estersTOP
Fatty acids of purified esters of AST are presented in Table 5. The FA profile of ester fractions of both samples contained the same predominant C18:1n-9 and C16:0 than in the complete
oil extracts, although minor FAs appeared in the EXK esters as caprylic C8:0, lauric C12:0, myristoleic C14:1n-7, and araquidic
C20:0, at low amounts (<3%). The predominance of C18:1n-9 and C16:0 in the esterified AST was similar to those obtained by
Wolfe et al. (1965), in O. rusticus. A conspicuous difference in the FA composition of AST esters between the two analyzed samples was that concerning the PUFAs
components. The AST esters of C. (C.) montezumae contained LA and ALA, while AST-esters of EXK contained noticeable enrichment of DHA in the form of monoester. This feature
was not observed in the EXK of O. rusticus, which displayed small amounts of LA and ALA and no DHA (Wolfe et al., 1965). Moreover, preferential accumulation of EPA and DHA as monoesters was determined in the marine red crab Pleuroncodes planipes (Coral-Hinostroza and Bjerkeng, 2002). To this regard, Vincent and Cecaldi (1988), stated that the unsaturation level of fatty acid esterifying carotenoids increases with the increasing level of molecule
oxygenation; thus, AST as one of the most oxygenated carotenoids would be expected to be most esterified with highly unsaturated
FAs. We think that during biosynthesis, the steric structure of DHA prevents the esterification of both sites of the carotenoid
giving preference to monoesterification. This particular association of DHA and EPA with AST has high pharmacological significance
because their antioxidant synergism at low concentrations has been demonstrated (Saw et al., 2013). The recovery of these valuable bioactive components from the EXK of P. (M.) bouvieri waste would improve the utilization and economic exploitation criteria of this crayfish.
Table 5. Astaxanthin esters and their fatty acid composition expressed as percentage (%) of the total
| Fatty acid |
C. (C.) montezumae whole animal
|
P. (M.) bouvieri EXK
|
| AST-diester |
AST-monoester |
AST-diester |
AST-monoester |
| SFA |
|
|
|
|
| C8:0 |
|
|
1.50 |
|
| C12:0 |
|
|
2.10 |
0.65 |
| C14:0 |
3.07 |
2.23 |
2.36 |
2.69 |
| C15:0 |
2.21 |
1.82 |
1.26 |
0.67 |
| C16:0 |
18.72 |
17.08 |
9.97 |
10.45 |
| C17:0 |
1.43 |
1.36 |
|
0.67 |
| C18:0 |
3.96 |
4.44 |
3.84 |
6.63 |
| C20:0 |
|
|
3.50 |
2.27 |
| C23:0 |
|
2.96 |
|
0.47 |
| MUFA |
|
|
|
|
| C14:1n-7 |
|
|
|
1.68 |
| C16:1n-7 |
2.47 |
1.21 |
1.40 |
0.81 |
| C17:1n-8 |
1.45 |
1.27 |
1.22 |
1.17 |
| C18:1n-9 |
16.62 |
17.0 |
14.92 |
2.97 |
| C22:1n-9 |
3.77 |
3.61 |
4.48 |
0.83 |
| PUFA |
|
|
|
|
| C18:2n-6 |
12.87 |
10.87 |
8.70 |
1.29 |
| C18:3n-3 |
6.02 |
4.54 |
4.10 |
0.53 |
| C20:2n-6 |
|
|
1.77 |
2.64 |
| C20:4n-6 |
|
|
|
0.49 |
| C20:5n-3 |
|
|
1.31 |
1.49 |
| C22:6n-3 |
|
|
7.30 |
30.46 |
| ΣSFA |
40.49 |
42.99 |
35.18 |
35.58 |
| ΣMUFA |
33.49 |
33.21 |
31.58 |
10.83 |
| ΣPUFA |
26.02 |
23.79 |
33.24 |
53.59 |
| EXK, exoskeleton. |
We conclude that C. (C.) montezumae and the TM of P. (M.) bouvieri are good sources of n-3 PUFAs of good nutritional and functional quality equivalent to some marine fish. Moreover, C. (C.) montezumae is a good source of dietary carotenoids, particularly of AST. The EXK of P. (M.) bouvieri represents a valuable potential source of n-3 PUFAs, free AST and the combined n-3 PUFAs-AST for different purposes (food
additives, animal feeding, or nutraceuticals). An increased accessibility of crayfish and their by-products through aquaculture
might open up new possibilities for obtaining this novel source of bioactive compounds.
ACKNOWLEDGEMENTSTOP
We thank MS Rosa Maria Castillo for her technical assistance in the laboratory. This research was supported by the Department
of Animal Nutrition, INCMNSZ, and partly, by Program 717, FUNSALUD. There was not any involvement of the supporting agencies
in the conduct of this work.
REFERENCESTOP
| ○ |
Ackefors H, Castell J, Örde-Öström IL. 1997. Preliminary results on the fatty acid composition of freshwater crayfish, Astacus astacus and Pacifastacus leniusculus, held in captivity. J. World Aquacult. Soc. 28, 97–105. http://dx.doi.org/10.1111/j.1749-7345.1997.tb00967.x |
| ○ |
Alvarez F, Villalobos JL, Armendáriz G, Hernandez C. 2012. Biogeographic relationship of freshwater crabs and crayfish along
the Mexican transition zone: reevaluating Rodríguez (1986) hypothesis. Rev. Mex. Biodivers. 83, 1073–1083.
|
| ○ |
AOAC. 2000. Official Methods of Analysis of AOAC International. Association of Official Analysis Chemists International. Washington,
D.C., USA.
|
| ○ |
Barros MP, Poppe SC, Bondan EF. 2014. Neuroprotective properties of the marine carotenoid astaxanthin and omega-3 fatty acids,
and perspectives for the natural combination of both in krill oil. Nutrients, 6, 1293–317. http://dx.doi.org/10.3390/nu6031293 |
| ○ |
Bosco AD, Castellini C, Bernardini M. 2001. Nutritional quality of rabbit meat as affected by cooking procedure and dietary
Vitamin E. J. Food Sci. 66, 1047–1051. http://dx.doi.org/10.1111/j.1365-2621.2001.tb08233.x |
| ○ |
Cerón-Ortiz AN, Moctezuma-Reséndiz O, Ángeles-Monroy MÁ, Montufar-Serrano E, León-Escamilla JA. 2015. Efecto interactivo del
alimento y la calidad de agua en el crecimiento y sobrevivencia de postlarvas de acocil de río Cambarellus montezumae. Rev. Mex. Biodivers. 86, 131–142. http://dx.doi.org/10.7550/rmb.48502 |
| ○ |
Cook CM, Hallaråker H, Sæbø PC, Innis SM, Kelley KM, Sanoshy KD, Berger A, Maki KC. 2016. Bioavailability of Long Chain Omega-3
Polyunsaturated Fatty Acids from Phospholipid-Rich Herring Roe Oil in Men and Women with Mildly Elevated Triacylglycerols.
Prostag. Leukotr. Ess. (PLEFA). In press. http://dx.doi.org/10.1016/j.plefa.2016.01.007 |
| ○ |
Coral-Hinostroza GN, Bjerkeng B. 2002. Astaxanthin from the red crab langostilla (Pleuroncodes planipes): optical R/S isomers and fatty acid moieties of astaxanthin esters. Comp. Biochem. Phys. B. 133, 437–444. http://dx.doi.org/10.1016/s1096-4959(02)00186-0 |
| ○ |
Cremades O, Parrado J, Alvarez-Ossorio MC, Jover M, de Teran LC, Gutierrez JF, Bautista J. 2003. Isolation and characterization
of carotenoproteins from crayfish (Procambarus clarkii). Food Chem. 82, 559–566. http://dx.doi.org/10.1016/s0308-8146(03)00011-6 |
| ○ |
Czeczuga B. 1971. Composition and tissue distribution of carotenoids and vitamin A in the crayfish Astacus leptodactylus (Esch.) (Crustacea, Decapoda). Comp. Biochem. Phys. B. 39, 945–953. http://dx.doi.org/10.1016/0305-0491(71)90118-0 |
| ○ |
Dose J, Matsugo S, Yokokawa H, Koshida Y, Okazaki S, Seidel U, Eggersdorfer M, Rimbach G, Esatbeyoglu T. 2016. Free Radical
Scavenging and Cellular Antioxidant Properties of Astaxanthin. Int. J. Mol. Sci. 17, 103. http://dx.doi.org/10.3390/ijms17010103 |
| ○ |
FAO. 2010. Fats and fatty acids in human nutrition. Report of an expert consultation.10–14 November 2008, Geneva. FAO Food
and Nutrition. Paper 91. Rome: Food and Agricultural Organisation of the United Nations.
|
| ○ |
Fernandes CE, da Silva Vasconcelos MA, de Almeida Ribeiro M, Sarubbo LA, Andrade SAC, de Melo Filho AB. 2014. Nutritional
and lipid profiles in marine fish species from Brazil. Food Chem. 160, 67–71. http://dx.doi.org/10.1016/j.foodchem.2014.03.055 |
| ○ |
García-Romero J, Ginés R, Izquierdo M, Robaina L. 2014. Marine and freshwater crab meals in diets for red porgy (Pagrus pagrus): Effect on fillet fatty acid profile and flesh quality parameters. Aquaculture, 420, 231–239.
|
| ○ |
Gonzalez-Baro MDR, Pollero RJ. 1988. Lipid characterization and distribution among 463 tissues of the freshwater crustacean
Macrobrachium borellii during an annual 464 cycle. Comp. Biochem. Phys. B. 91, 711–715. http://dx.doi.org/10.1016/0305-0491(88)90197-6 |
| ○ |
Harlioğlu AG, Aydin S, Yilmaz O. 2012. Fatty acid, cholesterol and fat-soluble vitamin composition of wild and captive freshwater
crayfish (Astacus leptodactylus). Food Sci. Technol. Int. 18, 93–100. http://dx.doi.org/10.1177/1082013211414261 |
| ○ |
Harlıoğlu MM, Köprücü K, Harlıoğlu AG, Yılmaz Ö, Yonar SM, Aydın S, Duran TÇ. 2015. Effects of dietary n-3 polyunsaturated
fatty acids on the nutritional quality of abdomen meat and hepatopancreas in a freshwater crayfish (Astacus leptodactylus). J. Food Compos. Anal. 41, 144–150. http://dx.doi.org/10.1016/j.jfca.2015.01.011 |
| ○ |
Hernández JA, Ochoa AA, Valerio-Alfaro G, Soto-Rodríguez I, Rodríguez-Estrada MT, García HS. 2014. Cholesterol oxidation and
astaxanthin degradation in shrimp during sun drying and storage. Food Chem. 145, 832–839. http://dx.doi.org/10.1016/j.foodchem.2013.08.098 |
| ○ |
Inoue T, Simpson KL, Tanaka Y, Sameshima M. 1988. Condensed astaxanthin of pigmented oil from crayfish carapace and its feeding
experiment. Nippon Suisan Gakk. 54, 103–106. http://dx.doi.org/10.2331/suisan.54.103 |
| ○ |
Konagai C, Yanagimoto K, Hayamizu K, Han L, Tsuji T, Koga Y. 2013. Effects of krill oil containing ω-3 polyunsaturated fatty
acids in phospholipid form on human brain function: a randomized controlled trial in healthy elderly volunteers. Clin. Interv. Aging. 8, 1247–1257. http://dx.doi.org/10.2147/cia.s50349 |
| ○ |
Meyers SP, Bligh D. 1981. Characterization of astaxanthin pigments from heat-processed crayfish waste. J. Agr. Food Chem. 29, 505–508. http://dx.doi.org/10.1021/jf00105a017 |
| ○ |
Ramírez-Silva I, Villalpando S, Moreno-Saracho JE, Bernal-Medina D. 2011. Fatty acids intake in the Mexican population. Results
of the National Nutrition Survey 2006. Nutr. Metab. 8, 33. http://dx.doi.org/10.1186/1743-7075-8-33 |
| ○ |
Ramprasath VR, Eyal I, Zchut S, Jones PJ. 2013. Enhanced increase of omega-3 index in healthy individuals with response to
4-week n-3 fatty acid supplementation from krill oil versus fish oil. Lipids Health Dis. 12, 1. http://dx.doi.org/10.1186/1476-511x-12-178 |
| ○ |
Sachindra NM, Bhaskar N, Mahendrakar NS. 2005. Carotenoids in different body components of Indian shrimps. J. Sci. Food Agric. 85, 167–172. http://dx.doi.org/10.1002/jsfa.1977 |
| ○ |
Sagi A, Rise M, Isam K, Arad S. (Malis). 1995. Carotenoids and their derivatives in organs of the maturing female crayfish
Cherax quadricarinatus. Comp. Biochem. Phys. B. 112, 309–313. http://dx.doi.org/10.1016/0305-0491(95)00069-0 |
| ○ |
Saw CLL, Yang AY, Guo Y, Kong ANT. 2013. Astaxanthin and omega-3 fatty acids individually and in combination protect against
oxidative stress via the Nrf2–ARE pathway. Food Chem. Toxicol. 62, 869–875. http://dx.doi.org/10.1016/j.fct.2013.10.023 |
| ○ |
Stanek M, Borejszo Z, Dąbrowski J, Janicki B. 2011. Fat and cholesterol content and fatty acid profiles in edible tissues
of spiny-cheek crayfish, Orconectes limosus (Raf.) from Lake Gopło (Poland). Arch. Pol. Fish. 19, 241–248. http://dx.doi.org/10.2478/v10086-011-0030-7 |
| ○ |
Ulbricht TLV, Southgate DAT. 1991. Coronary heart disease: seven dietary factors. The Lancet 338, 985–992. http://dx.doi.org/10.1016/0140-6736(91)91846-m |
| ○ |
Valfré F, Caprino F, Turchini GM. 2003. The health benefit of seafood. Vet. Res.Commun. 27, 507–512. http://dx.doi.org/10.1023/b:verc.0000014208.47984.8c |
| ○ |
Vincent M, Ceccaldi HJ. 1988. Relations entre acides gras et pigments caroténoïdes chez un crustacé copépode, Calanipeda aquae-dulcis. Biochem. Syst. Ecol. 16, 317–324. http://dx.doi.org/10.1016/0305-1978(88)90017-8 |
| ○ |
Wolfe DA, Rao PV, Cornwell DG. 1965. Studies on the fatty acid composition of crayfish lipids. J. Am. Oil Chem. Soc. 42, 633–637. http://dx.doi.org/10.1007/bf02541304 |
| ○ |
Zagalsky PF, Eliopoulos EE, Findlay JB, 1990. The architecture of invertebrate carotenoproteins. Comp. Biochem. Physiol. B 97, 1–18. http://dx.doi.org/10.1016/0305-0491(90)90171-o |